
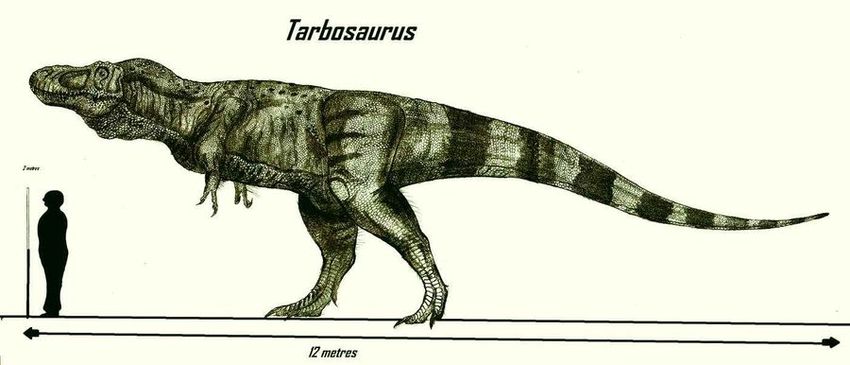
12-метровый Тираннозавр рекс, погибший в возрасте 28 лет. Музей Филда, Чикаго. Фото Александр Бабкин
Тираннозаврины – подсемейство вымерших ящеротазовых хищных рептилий входящее в семейство Тираннозаврид надсемейства Тираннозавроидов. Кроме того в подсемейство тираннозаврин, как предполагается, входит еще одна группа – Алиорамины.
Среди собственно тираннозаврин на сегодня известны 8 родов, каждый из которых представлен одним видом, кроме рода Дасплетозавров, состоящего из двух видов: торосус и хорнери. Остальные 7 видов это: Литронакс аргестес (Lythronax argestes), Тератофоней курри (Teratophoneus curriei), Жученгтиранн магнус (Zhuchengtyrannus magnus), Нанукзавр хоглунди (Nanuqsaurus hoglundi), Тарбозавр батаар (Tarbosaurus bataar) и Тираннозавр рекс (Tyrannosaurus rex).
Однако общепризнанной классификации тираннозавроидов не существует.
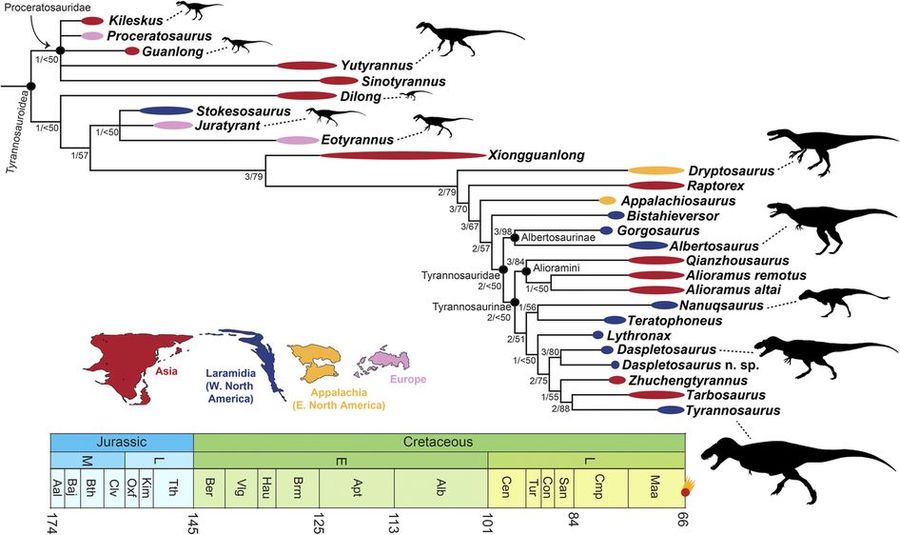
Тираннозаврины обитали на территории Северной Америки, Западной Европы, Восточной и Центральной Азии в конце мелового периода между около 80 млн лет и 66 млн лет назад. Предки тираннозаврин пришли в Северную Америку из Евразии в конце мела, когда образовался сухопутный мост через Берингов пролив.
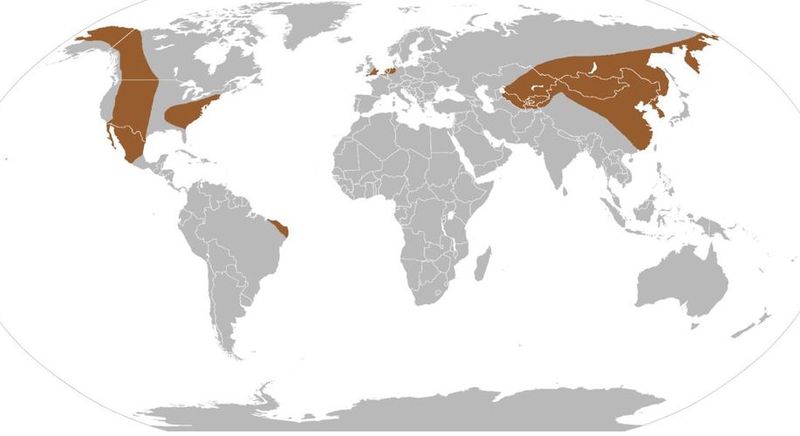
Останки тираннозавринов были найдены в различных экосистемах – приморских, удаленных от моря участках суши, болотистых местностях, засушливых и полузасушливых равнинах.
Тираннозаврины особенно характеризуются массивными головами, загнутыми бананоподобными зубами, миниатюрными передними лапами с двумя пальцами, структурой бедренных костей, мощным двойным килем и соединением носовых костей.
Нам известны только два тираннозавроида, обладавших перьями, но можно предположить, что все они были оперенными, в том числе и тираннозаврины. Есть идея, что молодняк тираннозавроидов был опушен, а взрослые особи утрачивали теплоизолирующий покров из-за перегрева. Однако полный перьевой покров ютираннуса, весившего больше тонны, противоречит этому аргументу. Возможно, что кроме оперения у тираннозаврин местами наличествовала и чешуя.
Размноженеие: Самцы-тираннозаврины должны были доставить сперму в клоаку самки. Это кажется невозможным, глядя на их строение, но множество рептилий и птиц спариваются без каких-либо действий, кроме прижимания клоак друг к другу. Тем не менее должно было быть нечто, что делало спаривание тираннозаврин более удобным. Этим нечто мог быть копулятивный орган похожий на пенис.
У тираннозавроидов многочисленные яйца формировались одновременно и откладывали они их сериями.
Литронакс аргестес – древнейший тираннозаврид – 80,6-80 млн лет. Юта. Длина – 7-8 м; рост – 2,5 м; масса – 2 тонны.

Литронакс
Дасплетозавры – 77-74 млн, род состоящий из двух видов — D. torosus, обнаруженный на территории канадской провинции Альберта и более поздний D. horneri, найденный в штате Монтана. Длина – до 9м; вес – до 3,8т.

Дасплетозавр. commons.wikimedia.org
Тератофоней курри – около 77-70,6 млн лет. Юта. Длина 5м.

Тератофоней. Автор NobuTamura
Жученгтиранн магнус – около 70 млн лет. Описание сделано по нижней челюсти и зубам, найденным в 2009г. строителями, которые готовили фундамент под палеонтологический музей. Находка сделана в провинции Шаньдун, восточный Китай. Предположительный рост ящера – 4м; длина – около 11м.
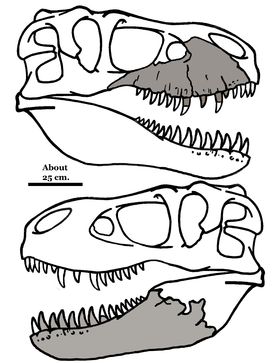
Закрашены сохранившиеся останки Жученгтиранна магнуса
Нанукзавр хоглунди – около 71-68 млн лет. Аляска. Длина до 6м.

Нанукзавр. Автор Nobu Tamura
Самыми известными в подсемействе являются тираннозавр и тарбозавр. Крупнейшие полные скелеты этих рептилий имеют длину 12м, высоту 4м, а оцениваемая масса – около 9т.
Тираннозавры жили 67-66 млн лет назад в западной части Северной Америке, которая в те времена представляла собой остров Ларамидию и были одним из последних видов ящеротазовых динозавров, существовавших перед астероидной катастрофой. Падение астероида стало заключительным аккордом продолжавшегося уже несколько миллионов лет вымирания сухопутной и морской фауны и флоры, вызванного климатическими изменениями и мощной вулканической деятельностью на полуострове Индостан (лавы плато Декан) накануне палеогенового периода.
Как и другие представители своего семейства, тираннозавр был двуногим хищником с массивным черепом уравновешенный длинным, жестким хвостом. По сравнению с большими задними лапами этого ящера его передние лапы были маленькими и имели два когтистых пальца. Тираннозавр охотился на гадрозавров, цератопсов, хотя некоторые исследователи предполагают, что питался он преимущественно падалью. Сейчас большинство специалистов считает, что тираннозавр мог как охотиться, так и питаться падалью.
Были найдены окаменевшие свидетельства агрессивных столкновений между тираннозавром и трицератопсом: следы от зубов на надбровном роге и некоторых костях шеи одного экземпляра трицератопса (впоследствии вместо сломанного рога вырос новый, а рана на шее зажила). В то же время, следы от зубов тираннозавров на костях трицератопсов без следов заживления достаточно распространены. Также обнаружены скелеты гадрозавров (утконосые динозавры) на костях которых имеются следы от зубов тираннозавров и тарбозавров, а у двух гадрозавров в костях ноги и хвоста были застрявшие зубы тираннозавра. 14% костей гадрозавров и 4% костей цератопсов извлеченных в Альберте (Канада) несут на себе следы зубов тираннозаврин. Содержимое кишечников и копролитов тираннозаврин также указывает на поедание различных гадрозавров.

Разные виды цератопсов

Череп Тираннозавра рекса. Музей истории природы, Берлин

Тираннозавр рекс в музее истории природы, Питтсбург, Огайо
Было найдено более 30 скелетов тираннозавров, некоторые из них сохранились почти полностью. Существуют фрагментарные окаменелости, принадлежавшие, возможно, еще более крупным тираннозаврам. Например, одна найденная 81-см верхнечелюстная кость могла принадлежать тираннозавру длиной 13,6м и массой в 12 тонн.
Шея тираннозавра была короткой и мускулистой, что позволяло ей удерживать массивную голову. Задние конечности были самыми длинными (по отношению к телу) среди всех хищных динозавров. Позвоночник состоял из 10 шейных, 12 грудных, 5 крестцовых и около 40 хвостовых позвонков. Тяжёлый и длинный хвост уравновешивал массивную голову и грузное туловище.
Самый большой из когда-либо найденных полных черепов тираннозавра в длину достигает примерно полутора метров. Большие окна в черепе способствовали уменьшению его веса и создавали необходимое пространство для прикрепления мышц. Его задняя часть была очень широкой, но рыло было узким, благодаря чему ящер обладал высокоразвитым бинокулярным зрением (способность видеть объекты в трехмерном виде). Бинокулярный диапазон у тираннозавра равнялся 55 градусам – больше чем у современного ястреба. Также было установлено, что острота зрения тираннозавра в 13 раз превышала таковую у человека, и, соответственно, превышала остроту зрения орла (которая в 3,6 раза выше человеческой) – это позволяло тираннозавру распознавать объекты на расстоянии 6км.
Носовая и некоторые другие кости были объединены. Они были воздухоносны. Конец верхней челюсти имел U-образную форму. Эти свойства увеличивали силы укуса.
У тираннозавра наличествовали нескольких типов зубов, различавшихся по форме и функциям. Зубы, располагающиеся на передней стороне верхней челюсти, плотно прилегают друг к другу, имеют D-образное поперечное сечение, долотообразное лезвие, укрепляющие гребни и изгиб внутрь. Эти особенности понижали риск слома зуба. Другие зубы более крепкие и массивные, по форме они больше напоминают бананы, чем кинжалы, они шире расставлены и тоже имеют укрепляющие гребни. Зубы верхней челюсти больше зубов нижней (кроме тех, что находятся в её задней части). Самый большой из найденных зубов вместе с корнем достигает высоты 30см.
Предположительно сила укуса тираннозавра была самой большой среди всех когда-либо живших наземных животных. Задние зубы взрослого T. rex могли смыкаться с силой 37 килоньютонов, что в 15 раз больше, чем наибольшая измеренная сила укуса современного рекордсмена – австралийского гребнистого крокодила. За один укус взрослый тираннозавр мог вырвать из тела жертвы кусок мяса весом в 70кг.
У T. rex быстрое увеличение массы начиналось с 14 лет (этому возрасту соответствует масса 1800кг). В течение этой стремительной фазы роста тираннозавр на протяжении 4 лет набирал по 600кг в год. Когда особь достигала 18-летнего возраста, её рост резко замедлялся.
Передние конечности тираннозавра весьма малы по отношению к общим размерам тела, в длину они достигают лишь одного метра, и некоторые учёные считают их рудиментарными органами. Но на их костях есть большие участки для прикрепления мышц, что говорит о значительной силе. Передние конечности могли служить T. rex для того, чтобы встать из позы отдыха. Предполагается также, что передние конечности могли использоваться для удержания полового партнёра во время спаривания или удержания жертвы, пока хищник убивал её своими огромными челюстями.
Кости передних конечностей T. rex имеют исключительно толстый поверхностный непористый слой, что позволяло им выдерживать большую нагрузку. Диапазон движений предплечья тираннозавра был ограничен, плечевые и локтевые суставы могли двигаться лишь на 40 и 45 градусов соответственно. Для сравнения: те же два сустава человека могут двигаться на 360 и 165 градусов соответственно.
В журнале Science в 2007г. сообщалось об исследованиях согласно которым коллаген, обнаруженный в очищенной кости T. rex, ближе всего к коллагену курицы и более далёк от коллагена лягушек и тритонов.
Многие учёные считают, что T. rex имел перья (по крайней мере, на отдельных частях тела), хотя прямых подтверждений этого нет. Это предположение основывается на наличии перьев у некоторых родственных видов меньшего размера – у некоторых больших тираннозавроидов нашли перья, покрывавшие большую часть тела.
Тираннозавр возможно поддерживал постоянную температуру тела, но это не означает, что он был полностью теплокровным. Такая терморегуляция может быть объяснена развитой формой мезотермии.
Тираннозавру рекс приписаны два отдельных отпечатка стопы и цепочка следов. Один из них и серия следов обнаружены в штате Нью-Мексико. След был оставлен в болотных угодьях. Длина отпечатка равняется 83см, ширина – 71см. Второй вероятный отпечаток стопы тираннозавра найден в штате Монтана. Длина этого следа составляет 72см.
Скорее всего, тираннозавр разворачивался медленно, вероятно, для поворота на 45° ему требовалась одна-две секунды. Причина этих сложностей заключается в том, что большая часть массы тираннозавра была удалена от его центра тяжести, однако он мог уменьшить это расстояние, выгибая спину и хвост и прижимая голову и конечности к туловищу. Предположительная оценка максимальной скорости тираннозавра – около 40 км/ч (11 м/с).
Тираннозавр обладал отличным обонянием. Вероятно, обоняние тираннозавра было сопоставимо с таковым у современных стервятников.
Ящер наиболее хорошо улавливал низкочастотные звуки.
Установлено, что тираннозавр имел самый большой относительный размер мозга среди всех нептичьих динозавров, кроме небольших представителей манирапторов. Относительный размер мозга тираннозавра лежит в пределах диапазона такового у современных рептилий, хотя и существенно выше среднего значения.
Во время изучения экземпляра Сью в Чикаго был обнаружен зуб другого тираннозавра, застрявший в шейных позвонках. У ювенильной особи тираннозавра Джейн зафиксированы впоследствии зажившие колотые раны на нижней челюсти и морде. Это может свидетельствовать об агрессивном поведении между тираннозаврами, но точно неизвестно, были ли тираннозавры активными каннибалами или просто участвовали во внутривидовой борьбе.
В Южной Дакоте была сделана находка скелетов трех тираннозавров в непосредственной близости друг от друга. Тираннозавры могли жить в стаях – размер мозга тираннозавра в три раза превышает обычный размер для животного таких габаритов, следовательно, ящер был способен к сложному социальному поведению. Тем не менее, гипотеза о стайной охоте тираннозавров подвергается жесткой критике со стороны других специалистов.
Тарбозавр батаар. Обнаруживаемые с 1946г. останки нескольких десятков особей Тарбозавр батаар, включая целые черепа и скелеты, позволяют воссоздать его внешний облик и сделать выводы об образе жизни, а также проследить его эволюцию. Тарбозавр обитал во времена позднего мела около 76-66 млн лет назад на территории нынешних Монголии, Китая и Казахстана.
Тарбозавры были крупными двуногими хищниками весом до 6 тонн, с непропорционально маленькими относительно остального тела двупалыми передними конечностями. В пасти тарбозавра располагались около 60 зубов до 85мм длиной. Хотя ряд исследователей считает тарбозавров падальщиками, более распространена точка зрения, согласно которой они были активными хищниками и занимали верхнее положение в экосистеме своей эпохи и региона, охотясь на крупных травоядных динозавров во влажных речных поймах.
По своему внешнему виду тарбозавр очень близок к тираннозавру.
В общей сложности обнаружены кости, принадлежащие более чем 30 экземплярам.

Скелет тарбозавра. Музей палеонтологии, Москва. Фото Александр Бабкин

Тарбозавр из Монголии
Хотя тарбозавры изучены меньше, чем тираннозаврины Северной Америки, доступного материала достаточно, чтобы учёные пришли к определённым выводам об их организме.
Немного уступая в размерах тираннозавру, тарбозавр всё равно оставался одним из самых крупных представителей семейства. Голова у тарбозавра была пропорционально крупнее по отношению к телу, чем у других тираннозаврин, включая тираннозавра. Самый крупный образец (длина черепа более 122см) был найден японско-монгольской экспедицией в 1984г. Масса тарбозавра считается такой же, или несколько меньшей, чем у тираннозавра сопоставимой длины.
Голова тарбозавра сидела на S-образной шее, а остальная часть позвоночника, включая длинный хвост, располагалась горизонтально. Крошечные передние конечности тарбозавра были ещё меньше по сравнению с остальным телом, чем у других представителей семейства. На каждой передней конечности, как и у других близкородственных видов, было по два когтистых пальца; у некоторых экземпляров имеется также третий палец, лишённый когтя. В отличие от передних, трёхпалые задние конечности тарбозавра были длинными и мощными. Длинный тяжёлый хвост служил противовесом голове и торсу, так что центр тяжести располагался в районе бёдер.
Череп высокий, как и у тираннозавра, но не такой широкий, особенно в затылочной части. То, что череп не расширяется к затылку, означает, что глаза тарбозавра не глядели прямо вперёд и что, возможно, в отличие от тираннозавра, он не обладал бинокулярным зрением. В челюстях размещалось 56-64 зуба, несколько больше, чем у тираннозавра, но не так много, как у тираннозаврин меньших размеров. Большинство зубов были овальными в сечении, за исключением полукруглых в сечении зубов, располагавшихся на предчелюстной кости. Самые длинные зубы располагались в верхней челюсти.

Фрагмент челюсти тарбозавра из Юго-восточного Казахстана. Saurian.blogspot

Фрагмент челюсти без зубов тарбозавра из Юго-восточного Казахстана. Музей природы, Алма-Ата. Фото Александр Бабкин
Ключевые отличия в строении черепа тарбозавра и североамериканских тираннозаврин, связаны с распределением давления по черепу в момент укуса. У североамериканских тираннозаврин основная часть нагрузки передавалась через верхнюю челюсть в спаянные носовые кости в верхней части морды. У тарбозавра эти костяные распорки отсутствовали. С другой стороны, у тарбозавра, в отличие от североамериканских тираннозаврин, была массивная, хорошо развитая задняя часть верхней челюсти. Ещё одним принципиальным различием между тарбозавром и североамериканскими тираннозавринами была жёсткая конструкция нижней челюсти. В то время как у тираннозаврин Северной Америки соединение задних и передних костей челюсти обладало известной гибкостью, нижнюю челюсть тарбозавра характеризует наличие запирающего механизма, образованного костной перемычкой между угловой костью и задней кромкой передней зубной кости.
Мозг тираннозаврин ближе по строению к мозгу крокодилов и других пресмыкающихся, чем к мозгу птиц. Общий объём мозга 12-метрового тарбозавра, по оценке исследователей, составлял только 184 см³. Большая обонятельная луковица и мощные обонятельный и концевой нервы свидетельствуют о хорошо развитом чувстве обоняния у тарбозавра, как и у тираннозавра. Большой слуховой нерв означает, что слух у тарбозавров был хорошим, позволяя вести звуковую коммуникацию и обеспечивать хорошую ориентацию в пространстве. Вестибулярный аппарат был хорошо развит – тарбозавры обладали хорошим чувством равновесия и координацией движений. С другой стороны, нервы и мозговые отделы, связанные со зрением, были маленькими и недоразвитыми. Крыша среднего мозга, отвечающая у пресмыкающихся за обработку зрительной информации, у тарбозавра была совсем маленькой, а зрительный и глазодвигательный нервы слабыми. Если у тираннозавра, с его широким черепом, глаза смотрели вперёд, обеспечивая бинокулярное зрение, то череп тарбозавра, как и у большинства других тираннозаврин, был у́же, и глаза располагались по разные стороны головы. Всё это позволяет сделать вывод о том, что основными чувствами, развитыми у тарбозавра, были обоняние и слух, но не зрение.
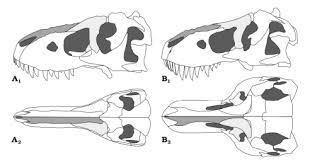
Череп тарбозавра слева и тираннозавра справа
В нэмэгэтских отложениях Монголии, где обнаружены скелеты тарбозавров, сохранились русла больших рек и почвенные наносы, свидетельствующие о том, что климат в это время был влажным. В формации попадаются окаменелые моллюски и останки других водных организмов, таких как рыбы и черепахи.
Взрослые тарбозавры, будучи крупнейшими хищными динозаврами в данной формации, вероятнее всего охотились на крупных гадрозавров, или на таких титанозавров, как немегтозавр. Вероятнее всего основными ландшафтами местообитания тарбозавров и тираннозавров были леса и лесостепные участки, где крупные хищники могли прятаться и устраивать засады, и где, согласно некоторым данным, предпочитали кормиться гадрозавры, а гадрозавры, как предполагается, были быстрее тираннозаврин на коротких дистанциях.
Тарбозаврам не могли составить конкуренцию более мелкие хищные динозавры (тероподы). Молодые особи тарбозавров, как это, вероятно, происходило с другими крупными тираннозавринами и происходит в настоящее время с крокодилами и комодскими варанами, возможно, занимали промежуточные экологические ниши между взрослыми тарбозаврами и более мелкими тероподами. Однако в 2010г. учёные из Китая и Японии пришли к выводу о падальческом образе жизни тарбозавра.

Гадрозавр. Южный Казахстан. Музей палеонтологии, Москва. Фото Александр Бабкин. Гадрозавры, вероятно, были основным источником пищи тарбозавров и тираннозавров
По материалам Википедии и книги Дэвида Хоуна «Хроники тираннозавра»
Гигантопитеки
Gigantopithecus – самый большой в истории примат из Южной и Юго-Восточной Азии. Гигантопитеки ответвились около 9 млн. лет назад от рамапитековых приматов. Известны три вида этого рода: Gigantopithecus bilaspurensis, Gigantopithecus blacki и Gigantopithecus giganteus. Вымерли в период между 300-100 тыс. лет назад.
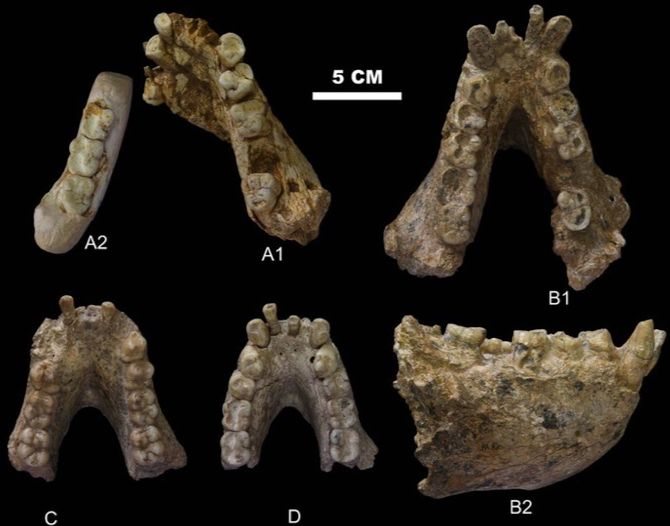
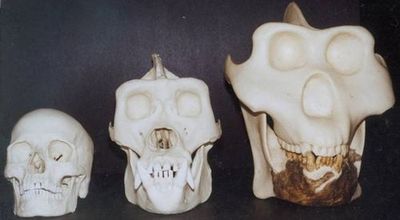
Останки гигантопитеков иногда встречаются в одних слоях с останками древних людей – Человек прямоходящий (Homo erectus) сталкивался с гигантопитеком, просуществовав с ним по соседству около 900 000 лет. Гигантопитеки известны почти исключительно по зубам, которых обнаружено уже несколько тысяч. Кроме зубов, найдено несколько обломков нижних челюстей и одна бедренная кость. Челюсти и зубы гигантопитеков были в полтора-два раза больше, чем у современных горилл – поэтому их рост вероятно доходил до 3-4 метра, а вес мог составлять 200–300 килограммов, что делает его вдвое крупнее африканской восточной гориллы. Гигантопитеки были специализированно-растительноядными приматами.
Просуществовав на протяжении почти 9 миллионов лет, гигантопитеки вымерли около 100 тысяч лет назад, когда климатические изменения привели к существенному похолоданию и уменьшению площади лесов, которые постепенно сменялись травянистыми равнинами. Другие растительноядные млекопитающие приспособились к жизни в саванне и питанию жесткой травой, а гигантопитек – исключительно лесной обитатель, не сумел адаптироваться и оказался изолирован в сохранившихся небольших участках леса.
Частичные нижние челюсти гигантопитека (Gigantopithecus blacki) из Китая: A1 – челюсть из пещеры Яньлянь; остальные фрагменты из пещеры Сяоянь |
Модели черепов человека, гориллы и гигантопитека |
 |
 |
Щелочные источники океанов – колыбель жизни?
На дне Атлантического океана существует тип гидротермальных источников, отличающийся от «черных» и «белых курильщиков» температурой и химическим составом извергаемой минеральной воды. Пока обнаружено только одно место, где расположены эти источники – в Атлантическом океане, в 15 км от Cрединно-Атлантического хребта в так называемом «Затерянном Городе», представляющим собой склон с башнеподобными белыми или зеленоватыми серпентинит-известняковыми образованиями высотой до 60м. Из этих башен выходят струи прозрачной воды с температурой около 200 гр.С насыщенные соединениями натрия, калия, кальция, магния, железа, марганца и кремния с кислородом, водородом и серой, а также обогащенные чистым элементарным водородом и углекислым газом. Башни Затерянного города состоят большей частью из известняка и серпентинита – сложнейшей метаморфической горной породы в состав которой входят до 70 минералов: минералы группы серпентина (соединения металлов с оксидом кремния и группами ОН), также кальцит, доломит, магнетит, хромит, макинавит и многие другие.
Серпентинитовые башни микропористы. Полости пор близки по размерам биологическим клеткам. Вот этим порам и приписывается роль инкубаторов, где химические процессы привели к возникновению живой субстанции, впоследствии вышедшей наружу в виде бактерий. Стенки пор очень тонкие и полупроницаемые, что весьма подходит для зарождения жизни в результате химических реакций при смешивании геотермальных вод с океанической водой.
Интересно, что такие щелочные источники были предсказаны Майком Расселом в 80-х годах после изучения им в Ирландии пузырчатых пород возрастом 350 млн. лет. Рассматривая в микроскоп сеть полостей породы, образованной древними щелочными источниками, Рассел пришел к идее о том, что минеральная химия в какой-то мере повторяет биохимические процессы, а может и наоборот. Интересно, что некоторые важные комплексы внутри бактерий идентичны некоторым минералам серпентинитовых башен по химическому составу и имеют очень близкое сходство по пространственному расположению атомов.
Рассел считал, что для инкубаторов жизни подходили бы источники с температурой не намного выше 100 гр.С и выделяющие свободный водород в насыщенной соединениями углерода водной среде. Такие условия, похоже, были в океане в архейское время 4-3 млрд лет назад. Океан, вероятно, был кислым из-за высокой концентрации углекислого газа в атмосфере, который растворялся в океане. На перегородках пор серпентинит-кальцитовых пород, слагающих башни щелочных источников, должны были образовываться градиенты концентраций веществ, что является одним из ключевых условий функционирования организмов. Большинство клеток организмов тратят большую часть своей энергии именно на поддержание подобных градиентов, для чего ими используются белки. По мнению Рассела – белки являются вторичной адаптацией организмов вышедших за пределы своих минералогических инкубаторов.
Однако щелочные источники никто прежде не наблюдал. Были известны только чёрные курильщики с температурой выше 400 гр.С. И вот в 2000 году американская экспедиция корабля «Атлантис» обнаружил именно такие щелочные источники в Атлантическом океане несколько в стороне от осевого рифта Срединно-Атлантического хребта.
Эти источники образуются не за счет вулканической энергии, а за счет выделения тепла при серпентинизации – метаморфического превращения базальтовых пород в серпентинит. Серпентинизация заключается в присоединении щелочных и щелочноземельных металлов и молекул воды к минералам базальта с выделением элементарного водорода. Это один из самых распространенных, если не самый распространенный геохимический процесс на Земле.
Жизнь в Затерянном городе идет за счет реакции свободного водорода с углекислым газом, которая проходит там прямо, тогда как на Земле эти реакции почти всегда осуществляются опосредованно через фотосинтез, что позволяет сначала отсоединить водород от воды.
Внутри пористых пород башен Затерянного города, открытого «Атлантисом», живут и процветают бактерии и археи. Археи – это, как и бактерии, одноклеточные безъядерные организмы, которые, однако имеют столько отличий от бактерий, что биологи склонны относить архей к отдельному царству. Мартин и Рассел выяснили, что механизмы синтеза белка у бактерий и архей весьма близки, а механизмы репликации ДНК отличаются радикально. Это можно было бы объяснить сильным эволюционным расхождением, но тогда становится не понятным, почему другие механизмы (например, транскрипция и трансляция) не стали столь же несхожими.
В 2003 году Мартин, Рассел и другой биохимик Евгений Кунин выдвинули гипотезу: общий предок бактерий и архей не был свободноживущим организмом, а был репликатором, живущим в минерале в неорганических клетках и жизнь из минеральных клеток выходила на свободу дважды – сначала в виде бактерии, а затем в виде архей. К такому выводу подталкивает и совершенно разные клеточные мембраны и стенки у бактерий и архей.
Башни Затерянного города действуют уже сорок тысяч лет – однако ожидать, что там продолжается абиогенное возникновение жизни (живое из неживого), пожалуй, не стоит. Химический состав океана сейчас уже далеко не тот, что был миллиарды лет назад – не хватает растворенного железа и, наверняка, еще каких-нибудь ингредиентов.
Но как жизнь вырвалась на свободу из башен гидротермальных источников? Рассел и другой американский ученый – Питер Митчелл предположили, что путь возникновения настоящих биологических клеток в виде бактерий лежал через формирование механизма хемиосмоса – биохимического механизма, с помощью которого осуществляется превращение энергии электротранспортной цепи в энергию аденозинтрифосфата. Аденозинтрифосфат (АТФ) выполняет важнейшую роль в обмене энергии и веществ в организмах и является универсальным источником энергии для всех биохимических процессов. Хемиосмотическая гипотеза Митчелла гласит, что электронпереносящие цепи митохондрий, хлоропластов и бактерий сопряжены с системой синтеза АТФ через разность электрохимических потенциалов протонов на сопрягающих мембранах.
Биохимический принцип это одно, а вот как физически образовалось клетка бактерии с ее мембраной не ясно.
Кроме того, совершенно не ясно, как безъядерные клетки бактерий и архей, то есть прокариоты, приобрели ядра, тем самым превратившись около 2 млрд лет назад в эукариоты – организмы, состоящие из ядерных клеток. Напомню, что грибы, растения и мы, животные, являемся эукариотами. Наличие или отсутствие ядра в клетке являет собой эволюционную пропасть. Преодоление этой пропасти вызывает столько споров в науке, что приходит мысль о сверхъестественном творении ядерной клетки. Был ли возможен эволюционный переход от прокариотов к эукариотам? Версий много, но отсутствуют данные могущие подтвердить или опровергнуть гипотезы.
Эволюционное древо показывает нам, что одноклеточные, а затем многоклеточные эукариоты возникли только однажды в промежутке, примерно, от 2 млрд до 1 млрд лет назад. Сложные формы жизни не эволюционировали из бактерий многократно и разновременно (растения от одних бактерий, животные – от вторых, грибы – от третьих). Ядерная клетка возникла из бактериальной только один раз.
Есть гипотеза о появлении некоего симбиоза бактерии и археи в результате чего возникла эукариотическая клетка. Но в течение минимум одного миллиарда лет эволюции такие симбиозы должны были неизбежно возникать многократно, если даже многие ранние эукариотные организмы и вымирали – ведь бактерии и археи здравствуют и доныне.
Это в полной мере относится и к появлению бактерий и архей. Жизнь вышла из щелочных источников океана только дважды в виде бактерий и архей? И это за 1,5 миллиарда лет существования архейского океана с подходящим химическим составом?
Создается впечатление, что был, а может и существует до настоящего времени внешний контроль над появлением жизни и ее эволюцией. Учитывая, что наука так и не смогла создать искусственный организм и, похоже, никогда и не создаст, приходишь к выводу: да – возможно жизнь и появилась в щелочных источниках океана, однако не самопроизвольно.
Известняково-серпентинитовая башня в Затерянном городе. Фото с "Элвина" |
|---|

Членистоногие появились за несколько миллионов лет до кембрия
Международная группа ученых проделала объемную и очень тонкую работу по оценке темпов эволюции членистоногих, этого самого разнообразного типа животных (только современных видов насчитывается более 10 млн), во временной окрестности нижней границы кембрийского периода. Сделан вывод, что в начале кембрия у членистоногих действительно произошло резкое (в среднем в 4–5 раз) повышение скорости эволюции по сравнению со значениями, обычными для этого типа в дальнейшем. Из выстроенной модели следует, что основные ветви членистоногих возникли за очень небольшой по геологическим меркам отрезок времени, как раз охватывающий границу докембрия и кембрия. Предполагается, что долгой докембрийской истории членистоногие (именно членистоногие, а не их отдаленные родственники) не имели: они появились всего за несколько миллионов лет до конца венда, и вряд ли раньше.
В начале кембрия произошло важнейшее эволюционное событие, которое называют кембрийским взрывом, — массовое появление в палеонтологической летописи множества современных типов животных, остатков которых в более древних слоях нет.
В кембрийском периоде (541млн– 485 млн лет назад) появляются все типы животных обитающие в последующей истории, кроме одноклеточных (с рифея), кольчатых червей (с рифея), губок (с венда), медузовидных стрекающих (с венда), плоских червей (с венда), иглокожих (с венда) и членистоногих (с конца венда), которые известны из отложений, образованных в последние несколько миллионов лет вендского (эдиакарского) периода протерозойского эона, примерно от 545 млн лет назад до начала кембрия 541 млн назад.
Гипотеза «долгой скрытой докембрийской эволюции» многоклеточных животных довольно сильно поддерживается молекулярной филогенетикой. Попросту говоря, белки и гены разных современных типов животных зачастую отличаются друг от друга таким количеством замен (аминокислотных и нуклеотидных), которое «всего лишь» за 540 миллионов лет накопиться не могло. Если, конечно, не предполагать, что когда-то в прошлом имело место резкое ускорение эволюции.
В 2011 году известный американский палеонтолог Дуглас Эрвин опубликовал аналитический обзор современного состояния проблемы происхождения типов животных, в котором постарался учесть как палеонтологические, так и молекулярно-генетические данные. Эрвин пришел к выводу, что первые многоклеточные животные появились примерно 800 млн лет назад, а почти все современные типы животных возникли в течение эдиакарского периода, который начался 635 млн лет назад и закончился 541 млн лет назад. Эдиакарий, который в нашей литературе гораздо чаще называют вендом, — это последний период протерозоя (2500 млн лет - 541 млн лет назад). То есть у Эрвина получается, что «скрытая докембрийская эволюция» действительно была. Просто в начале кембрия животные увеличили размер, приобрели скелеты, и поэтому их остатки стали гораздо доступнее палеонтологам. А эпохой взрывного возникновения новых эволюционных ветвей начало кембрия не являлось.
В 2013 году несколько ученых решили еще более тщательно исследовать проблему реальности кембрийского взрыва, сосредоточившись на данных по единственному типу, взятому в качестве примера, — по членистоногим. За эту работу взялись два биолога из австралийского города Аделаида — палеонтолог Майкл Ли (Michael Lee) и генетик Жюльен Субриер (Julien Soubrier), к которым присоединился известнейший специалист по эволюции членистоногих англичанин Грегори Эджкомб (Gregory Edgecombe).
Членистоногие были выбраны по четырем причинам. Во-первых, эта группа животных невероятно многочисленна и разнообразна, и она была такой уже в кембрии. Во-вторых, членистоногие обладают весьма сложным строением, а это значит, что исследователи получают много признаков для анализа. В-третьих, тело членистоногих покрыто хитиновым наружным скелетом, поэтому его части сохраняются в ископаемом состоянии лучше, чем остатки большинства других типов животных. И, в-четвертых, эволюция членистоногих хорошо изучена: для них построено подробное и достаточно надежное эволюционное древо, на которое можно опереться.
Ли, Субриер и Эджкомб исходили из того, что последовательность ветвлений этого древа в основном уже установлена. Их интересовали исключительно датировки точек дивергенции — развилок древа.
Исследователи с гордостью пишут, что набор данных, с которыми они работали, был довольно обширным: 395 морфологических признаков и последовательности 62 ядерных генов. По всему этому массиву было построено единое древо с использованием байесовского метода расслабленных часов (Bayesian relaxed-clock method). Байесовским методом называется способ построения эволюционных деревьев, использующий ряд формул из теории вероятностей , где скорость эволюции не предполагается постоянной. Ли, Субриер и Эджкомб приняли меры к тому, чтобы используемые ими программы анализировали морфологические признаки наравне с молекулярными. В этом отношении их исследование принципиально отличается от многих современных работ, где деревья строятся по молекулярным последовательностям, а с морфологическими данными только сопоставляются постфактум.
Главный вывод, сделанный исследователями, состоит в том, что эволюция членистоногих в начале кембрия действительно была очень быстрой. Для морфологических признаков раннекембрийская скорость эволюции превосходит среднюю по фанерозою (эон после протерозоя) примерно в 4 раза, для молекулярных — примерно в 5,5 раз. В отдельных ветвях, на коротких отрезках времени раннекембрийская скорость эволюции превосходит среднефанерозойскую даже в 16 раз. При этом скорости молекулярной и морфологической эволюции, оцениваемые отдельно, хорошо коррелируют друг с другом; это означает, что морфология является достаточно надежным показателем скорости генетических изменений, и наоборот.
Ничего нереального в таких скоростях эволюции нет. Никаких механизмов, неизвестных современной эволюционной теории, здесь постулировать не приходится.
Ли, Субриер и Эджкомб считают, что группа Panarthropoda, в которую вместе с членистоногими входят еще два типа — онихофоры и тихоходки — возникла все-таки до начала кембрия, но не раньше 650 миллионов лет назад. Что касается самих членистоногих, то их главная дивергенция произошла фактически в самый момент начала кембрия. На итоговом древе видно, что расхождение ветвей Chelicerata (мечехвосты и паукообразные) и Mandibulata (многоножки, ракообразные и насекомые) произошло 543 миллиона лет назад, а расхождение многоножек и ракообразных — 538 миллионов лет назад (рис. 3). Получается, что Chelicerata и Mandibulata разошлись перед самым началом кембрия, а многоножки и ракообразные — сразу после него. Понятно, что точность таких датировок не абсолютна. Но они явно указывают, что рождение главных ветвей членистоногих произошло в ближайшей временной окрестности нижней границы кембрия. На этот момент действительно пришелся пик изменений.
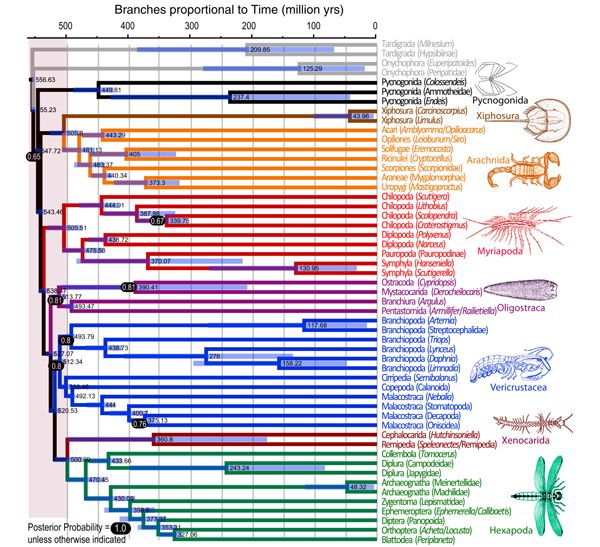
Когда жило самое первое членистоногое, сказать, конечно, пока точно нельзя, но если исходить из обсуждаемой модели — то примерно 550 миллионов лет назад (не раньше 556 и не позже 547 миллионов лет назад).
Докембрийская эволюция у членистоногих была, но была она очень короткой — буквально последние несколько миллионов лет перед границей кембрия.
Ученые подчеркивают, что даже если «удревнить» происхождение членистоногих, погрузив его еще на несколько десятков миллионов лет в докембрий, вывод о резком ускорении эволюции в начале кембрия все равно сохранит силу. Предположение, что раннекембрийская скорость эволюции была равна среднефанерозойской, потребует вывода, что Panarthropoda, например, возникли 940 миллионов лет назад — а это уж совсем малоправдоподобно.
Очень соблазнительно предположить, что интервал между датами 545 и 535 миллионов лет назад и есть тот момент времени, когда членистоногие возникли. Можно добавить, что первые представители современных групп ракообразных появляются в палеонтологической летописи 510 миллионов лет назад. Отрезка времени примерно в сорок миллионов лет, видимо, вполне достаточно не только для того, чтобы новый тип возник, но и для того, чтобы его представители набрали разнообразие и приобрели устойчивый набор признаков; кембрийские ракообразные отличаются от современных не так уж и сильно.
Поздний венд. Отпечаток членистоногого Parvancorina sagitta. Берег Белого моря |

Поздний венд. Xenusion auerswaldae. Германия |
Поздний венд. Трилобитообразное. Берег Белого моря |
|---|


Источник: Michael S. Y. Lee, Julien Soubrier, Gregory D. Edgecombe. Rates of Phenotypic and Genomic Evolution during the Cambrian Explosion // Current Biology. 2013. V. 23. P. 1889–1895
и Сергей Ястребов «Членистоногие подтверждают реальность кембрийского взрыва» Elementy.ru 17.11.2013: https://elementy.ru/novosti_nauki/432133/Chlenistonogie_podtverzhdayut_realnost_kembriyskogo_vzryva
Палеогеновые аммониты
Еще 7 лет назад в издании Bulletin of the Geological Society of Denmark вышла поистине
сенсационная публикация, посвященная обнаружению многочисленных раковин
аммонитов Baculites vertebralis и Hoploscaphites constrictus в отложениях,
возраст которых датировался датским веком палеогена. Сенсационность этой работы
заключалась в том, что до неё никому не удавалось обнаружить раковины аммонитов
в слоях, сформировавшихся после окончания мезозойской эры и лежащих выше
границы мелового и палеогенового периодов. То есть такие раковины в
палеогеновых слоях находили, но они явно были переотложенными – вымытыми
морским приливом из более древних отложений и снова захороненными вперемежку с более молодыми кайнозойскими раковинами. Но обнаруженные на территории Дании в отложениях датского же века палеогена аммониты переотложенными не были – это были раковины хорошей сохранности, заполнявшая их порода ничем (ни внешне, ни по изотопному составу, ни по включениям микрофоссилий) не отличалась от вмещающей породы,
датский возраст которой у авторов публикации не вызывал никаких сомнений. Как ни
странно, эта публикация не вызвала особого оживления в прессе (все-таки
аммониты не динозавры) и осталась малоизвестной за пределами научного
сообщества, но палеонтологов она заинтересовала.
И вот в 2012 году в журнале
Acta Palaeontologica Polonica вышли сразу четыре публикации, посвященные
вымиранию аммонитов на границе мела и палеогена. Авторы этих публикаций независимо друг от друга изучили изменение аммонитовых фаун в нескольких местонахождениях в различных точках земного шара: на побережье
Тихого океана (Россия), в штате Нью-Джерси (США), в горах Копетдаг
(Туркменистан) и в нескольких государствах Южной Америки.
Наибольшее
количество пограничных мел-палеогеновых разрезов было изучено в Южной Америке. Авторы этой работы искали аммонитов в
верхнемаастрихтских отложениях на территории Мексики, Колумбии, Бразилии, Чили
и Аргентины. Оказалось, что в целом вымирание аммонитов в морях, существовавших на территории современной Южной Америки, началось задолго до конца маастрихтского века. В первую очередь аммониты начали исчезать в экваториальных и тропических районах, а дольше всего они продержались в высоких широтах, в бореальном поясе. Тем не менее, и в южных районах Латинской Америки аммониты исчезают до окончания маастрихтского века.
Очень похожие результаты были получены при изучении позднемеловых слоев на российском Дальнем Востоке, на побережье Тихого океана. Последние аммониты в этих краях, представители родов Zelandites и Gaudryceras, встречаются в двух метрах ниже мел-палеогеновой границы. По мнению автора публикации, это означает, что аммониты исчезли здесь примерно за 20 тысяч лет до конца мезозоя.
Таким образом,
оказалось, что в двух регионах Земного шара аммониты просто не дожили до падения метеорита, который, как считается, положил конец эпохе динозавров.
Однако изучение верхнемеловых слоев в двух других местонахождениях принесло
совершенно иные результаты. В отложениях верхнего мела на реке Сумбар в Туркменистане палеонтологами были обнаружены раковины двух видов аммонитов: Baculites cf.vertebralis и Hoploscaphites constrictus johnjagti. В этих местах маастрихтские мергели (верхний мел) и датские мергели и известняки (нижний палеоген) разделяет тонкая
шестисантиметровая прослойка глины с повышенным содержанием иридия. Эта
иридиевая аномалия обнаружена и изучена во многих регионах Земли. Иридий редко
встречается в осадочных породах, в больших количествах он присутствует только в
метеоритах и его повышенное содержание в пограничном слое связано, по-видимому, с испарением части метеоритного вещества при падении на Землю крупного космического тела. Изучение горизонтов, лежащих ниже этой прослойки, показало, что аммониты реки Сумбар, в отличие от своих южноамериканских и дальневосточных родственников вовсе не собирались вымирать. Раковины аммонитов встречаются вплоть да самой верхней границы маастрихтских отложений, а один Hoploscaphites был найден выше иридиевой аномалии – в мергелях нижнего палеогена. Авторы исследования отмечают, что эта единичная находка не может служить доказательством того, что аммониты в этом месте дожили до палеогена, возможно, что раковина была вымыта из маастрихтских слоев и переотложена спустя многие годы после смерти аммонита. Однако, не вызывает сомнений тот факт, что на территории современного Туркменистана аммониты жили и неплохо себя чувствовали вплоть до самого конца мелового периода.
Еще более любопытная и странная картина открылась глазам исследователей в штате
Нью-Джерси. Там тоже была обнаружена иридиевая аномалия – слой с повышенным содержанием иридия. Но аммониты в этом местонахождении были не только под, но и над этим слоем! Причем выше лежали аммониты хорошей сохранности с неповрежденными раковинами, однозначно не переотложенные, не подвергавшиеся никакому размыву. Авторы публикации отмечают, что выше иридиевой аномалии можно выделить два слоя с аммонитами. Нижний слой имеет мощность 20 см и отличается большим количеством раковин двустворчатых моллюсков рода Pinna и хорошей сохранностью многочисленных аммонитов.
В общем - некоторые аммониты пережили меловой период и это вполне убедительные открытия. А сомнения в этом выглядят натянутыми.

Один из последних аммонитов. Фото из статьи "Terminal Maastrichtian ammonites from Turkmenistan, Central Asia" в журнале Acta Palaeontologica Polonica
Киин Кериш
В урочище Киин Кериш к северу от озера Зайсан представлены великолепные обнажения глин, алевритов и песчаников эоценовой и олигоценовой эпох палеогена (40-25 млн.лет). Обнаруженные здесь отпечатки листьев древесной палеофлоры подробно представляют смену субтропической эоценовой на теплоумеренную листопадную олигоценовую флору. Смена типов леса на Зайсанской равнине у берегов Джунгарского озера в палеогеновом периоде маркирует похолодание климата планеты вызванное формированием ледникового панциря Антарктиды в начале олигоцена.
Белые и желтые глины олигоцена и красные глины эоцена |
 |
|---|---|
 |
 |
 |
 |
Останец "Кораблик" содержащий множество отпечатков листьев теплоумеренной Тургайской флоры раннего олигоцена |
 |
 |
 |


Строматолиты - древнейшая форма жизни
Строматолиты - это рифовые тонкослойчатые столбики или холмики различной формы, состоящие из карбоната кальция и песчано-глинистого материала. Строматолит образуется в результате жизнедеятельности сообщества бактерий, называемого цианобактериальным матом (цианобактерии еще называются сине-зелеными водорослями). Маты существуют во многих районах мира, однако в современное время настоящие строматолиты существуют только в Акульем заливе на западном побережье Австралии, на атлантическом побережье Багамских островов и в озере Унтерзее в Антарктиде. Строматолиты достоверно появляются в геологической летописи в древнейших осадочных формациях Уарравуна (Западная Австралия) возрастом в 3,5 млрд лет – это древнейшая известная форма жизни. Наибольший расцвет цианобактерий пришелся на протерозойскй эон, затем их роль резко снизилась. Цианобактерии были и остаются самой распространенной группой организмов на планете.
Цианобактерии выделяют свободный кислород, одновременно химически связывая водород и углерод. Цианобактерии замечательны тем, что способны использовать атмосферный азот и превращать его в органические формы азота. При фотосинтезе они могут использовать углекислый газ как единственный источник углерода. В отличие от фотосинтезирующих бактерий, цианобактерии при фотосинтезе выделяют молекулярный кислород. В течении прошедших 3-х миллиардов лет до начала кембрия они являлись основным, наряду с фотохимическими реакциями в верхних слоях атмосферы, источником свободного кислорода в атмосфере Земли.
Цианобактерии относятся к подцарству оксифотобактерии. К ним относятся цианобактерии и хлороксибактерии. Эти бактерии имеют одиночные и колониальные формы. Колонии создают органогенные известковые постройки (строматолиты). В периферической части клеток цианобактерий диффузно распределены синий и бурый пигменты, определяющие в сочетании с хлорофиллом сине-зеленый цвет этих организмов. Некоторые цианобактерии могут иметь дополнительные пигменты, изменяющие их характерный цвет до черного, коричневого, красного. Цвет Красного моря определяется широким распространением в нем пурпурно пигментированных сине-зеленых. Цианобактерии могут использовать как солнечную энергию (автотрофность), так и энергию, выделяющуюся при расщеплении готовых органических веществ (гетеротрофность). Размножаются они только бесполым путем.
Мат, располагающийся на верхней поверхности создаваемого строматолита, представляет собой плотный многослойный "ковер" общей толщиной до 2 см; основу его составляют нитчатые либо пальмеллоидные цианобактерии, однако помимо них в формировании сообщества участвуют и другие бактерии. Внутри мата легко различимы несколько слоев:
1. Плотный верхний слой - поверхность роста (1-1,5 мм), в котором есть продуцирующие кислород автотрофы - цианобактерии-фотосинтетики, и гетеротрофы - облигатно аэробные (неспособные жить в отсутствие кислорода) бактерии.
2. Тонкая (менее 1 мм) подкладка; в ней автотрофами служат бактерии, осуществляющие бескислородный фотосинтез (при котором источником водорода является не вода, а другие вещества, например, сероводород - тогда выделяется не молекулярный кислород, а сера). Гетеротрофами же служат факультативные аэробы, которые могут использовать кислород, но в его отсутствие способны удовлетворять свои потребности в энергии не дыханием, а брожением.
3. Ниже двух верхних слоев, в которых идут процессы фотосинтеза и которые в совокупности называют фотической зоной мата, лежит толстая бескислородная зона, в которой процветают разнообразные анаэробы: для них молекулярный кислород не нужен или даже является ядом.
На поверхность обитающего на мелководье мата постоянно выпадают частички осадка (обычно кристаллы карбоната кальция), которые затемняют соответствующие участки фотической зоны и должны постепенно ухудшать условия происходящего в ней фотосинтеза - вплоть до неизбежной гибели фотосинтетиков. От катастрофы эти микроорганизмы спасает положительный фототаксис: по мере того как интенсивность света в фотической зоне падает, они мигрируют вверх сквозь накапливающийся слой осадка, образуя выше него новую поверхность роста с подкладкой. Миграция эта идет разными способами: нитчатые формы обладают способностью к скользящему движению сквозь осадок, пальмеллоидные - к ускоренному нарастанию верхних частей колонии. Оказавшиеся таким образом внутри бескислородной зоны мата осадки структурируются и слой за слоем наращивают верхнюю поверхность строматолита; измерения показали, что современные строматолиты прирастают со скоростью около 0,3 мм/год. Каждый слой в свою очередь состоит из пары слоев светлого и темного, состоящих из соответственно более мелких и более крупных кристаллов карбоната кальция; крупные кристаллы образуются в тот сезон года, когда процесс осаждения идет быстрее.
Подчеркнем: вещество, из которого построен строматолит, не создается матом; последний лишь структурирует естественное осадконакопление. Строматолиты чаще всего состоят из карбоната кальция потому лишь, что карбонатный тип осадконакопления в море наиболее обычен, однако в иных гидрохимических условиях формируются строматолиты фосфатные, кремнеземовые, железистые и пр. Многослойная расцветка строматолитов может меняться в течении суток, поскольку обитатели нижних слоев могут подниматься в темное время наверх и наоборот. Скользят бактерии вверх и вниз со скоростью до 2см в час.
Не все маты представляли собою донные сообщества. В онколитах - более или менее сферических строматолитах, известковые слои располагаются не линейно, а образуют концентрическую структуру. Предполагается, что онколиты создавались особыми бактериальными сообществами, имевшими облик плавающих в толще воды шаров, оболочка которых представляла собой мат; осадки отлагались внутри от оболочки, а плавучесть (вероятно только в начальной стадии роста) ему придавали образуемые при метаболизме газы, не способные покинуть замкнутую полость внутри колонии.
Мат представляет собой высокоинтегрированное сообщество с чрезвычайно сложно организованной структурой, два верхних слоя составлены продуцентами и консументами первого порядка ("растениями" и "травоядными"), соединенными в пищевую цепь пастбищного типа, а нижний слой сформирован редуцентами ("падальщиками"), получающими органику из верхних слоев. Более того, измерения показали, что мат является вообще одной из самых сбалансированных экосистем: он производит ровно столько органики и кислорода, сколько тут же расходует в процессе своей. Именно такая структура "производства и потребления" в прошлом позволяла матам образовывать кислородные оазисы и постепенно увеличивать их, вместо того чтобы пытаться сходу изменить всю эту бескислородную среду. Фотосинтезирующие бактерии подкладки имеют пигменты, позволяющие им поглощать свет из иной части спектра, чем их "коллеги" - цианобактерии из вышележащего слоя.
Таким образом, поверхность роста становится для жителей подкладки как бы прозрачной, что позволяет утилизовать всю поступающую в фотическую зону световую энергию. Более того, поскольку солнечный спектр меняется в течение дня ("покраснение" Солнца на восходе и закате), для улучшения использования энергии в мате происходят упорядоченные вертикальные миграции - смена горизонтов, занимаемых бактериями с разными типами пигментов.
Строматолиты обитали в соленых и пресных водах. В протерозое из строматолитов состояли огромные рифы мощностью в сотни метров. Отдельные глубоководные строматолиты достигали высоты 75м. Протерозойские строматолиты достигли высокого уровня сложности: появились формы со всевозможными ветвящимися столбиками, козырьками, разнообразной слоистостью и микроструктурой и т. п. Современные строматолиты, образуемые бактериальными матами, устроены намного проще.
По материалам Medbiol.ru
Онколит. Южный Казахстан |
Строматолит возрастом 3,5 млрд лет. Западная Австралия. www.Сarleton |
|---|---|
Строматолиты в заливе Шак (Акулий) в Западной Австралии. 100travels.ru |
Современные строматолиты высотой до 0,5м в озере Унтерзее. Антарктида. Фото с видео ВВС |
Бактерии из строматолитов. 775 млн. лет. Рифей. Южный Казахстан. Фото J. William Schopf |
Гигантские строматолиты. all-geo.org |
Протерозойские строматолиты. Плато Анабар. Сибирь. Фото Виталий Горшков |
Разрез строматолита. Фото James St. John |







Окаменелые деревья
Окамене́лое де́рево – древесина деревьев, произраставших в прошедшие геологические эпохи, замещённая различными минералами кремнезёма (халцедоном, кварцем или аморфным опалом). При этом структура дерева нередко хорошо сохраняется. Процесс окаменения происходит под землёй, когда дерево оказывается похороненным под осадком, но не портится из-за недостатка кислорода. Богатая минеральными веществами вода, текущая сквозь осадок, приносит минералы в клетки растения, и когда лигнин и целлюлоза распадаются, остаётся камень, повторяющий первоначальную форму. При этом все органические вещества замещаются минералами, а оригинальная микроскопическая структура дерева сохраняется. Наличие тех или иных примесей химических элементов, таких как железо, марганец, медь, во время петрификации (процесса окаменения) придает создающемуся окаменелому дереву ту или иную окраску из довольно широкого диапазона цветов.
Секвойя. Меловой период. Казахстан. Долина Или |
Окаменелая древесина секвойи. Меловой период. Казахстан. Долина Или |
|---|---|
Годовые кольца ствола диаметром 70см. Узбекистан. Музей геологии Узбекистана |
Палеоген. Южный Казахстан. Музей природы Академии наук Казахстана. Алматы |
Остров Лесбос. Греция. Фото Wald Riesenbaum |
|
Национальный парк Окаменелый лес. Аризона. Триас. nps.gov |
|
Национальный парк Окаменелый лес. Аризона. Триас. Мtritter.org |
Национальный парк Окаменелый лес. Аризона. Триас. Фото Stefan Pauli |
Национальный парк Окаменелый лес. Аризона. Триас. pikabu.ru |
Национальный парк Окаменелый лес. Аризона. Триас. Отполированный спил. www.geoclassica |
Аргентина. Фото Alberto Azparren |
Нью-Мексико. Аmericansouthwest.net |











